Applications des champs magnétiques (MF = champ magnétique) : un aperçu
Auteurs : Mohamed Sarraf / Sunita Kataria / Houda Taimourya / Lucielen Oliveira Santos / Renata Diane Menegatti / Meeta Jain / Muhammad Ihtisham / Shiliang Liu
- Collège d’architecture paysagère, Université agricole du Sichuan, Chengdu 611130, Chine
- Département des sciences horticoles, branche de Shiraz, Université islamique d’Azad, Shiraz 71987-74731, Iran
- École de biochimie, Devi Ahilya Vishwavidyalaya, Indore 452001, Inde
- Département d’Horticulture, Complexe Horticol d’Agadir (CHA), Institut Agronomique et Vétérinaire Hassan II, Agadir 80000, Maroc
- Laboratoire de biotechnologie, École de chimie et alimentation, Université fédérale de Rio Grande, Rio Grande-RS 96203-900, Brésil
- Département de botanique, Institut de biologie, Université fédérale de Pelotas, Rio Grande-RS 96203-900, Brésil
- Collège d’horticulture et de foresterie, Université agricole de Huazhong, Wuhan 430070, Chine
*
Auteurs auxquels la correspondance doit être adressée.
Plantes 2020 , 9 (9), 1139 ; https://doi.org/10.3390/plants9091139
Publié: 3 septembre 2020
Extraits (MF = champ magnétique)
Le rendement des cultures peut être augmenté par l’établissement d’un peuplement végétal adéquat en utilisant des graines à taux de germination et vigueur élevés. Divers traitements de pré-semis sont adoptés pour atteindre cet objectif. L’une de ces approches est l’exposition des semences à un champ magnétique (MF) de niveau faible à moyen, en modes pulsé et continu, car elles ont montré des résultats positifs dans un certain nombre de semences de cultures.
Sur la base de la sensibilité des plantes au MF, différents types de MF ont été utilisés pour les études de magnéto-amorçage, tels que les champs magnétiques homogènes statiques faibles (0-100 μT, y compris le GMF), les champs magnétiques homogènes forts (milliTesla à Tesla), et champs magnétiques à extrêmement basse fréquence (ELF) de densités de flux magnétique faibles à modérées (plusieurs centaines de μT).
L’application agronomique des FM dans les plantes a montré un potentiel pour modifier les systèmes de production végétale conventionnels ; augmenter les taux moyens de germination et la croissance des racines et des pousses ; avoir une productivité élevée ; augmenter la teneur en pigments photosynthétiques ; et intensifiant la division cellulaire, ainsi que l’absorption d’eau et de nutriments. En outre, différentes études suggèrent que les MF préviennent les dommages importants produits/infligés par les maladies et les ravageurs sur les cultures agricoles et d’autres plantes économiquement importantes et aident à réduire les dommages oxydatifs causés aux plantes par des situations de stress.
Une meilleure compréhension des interactions entre le MF et les réponses des plantes pourrait révolutionner la production agricole grâce à une résistance accrue aux maladies et aux conditions de stress, ainsi qu’à la supériorité de l’utilisation des nutriments et de l’eau, entraînant une amélioration du rendement des cultures. Dans cette revue, nous résumons les applications potentielles de MF et les processus clés impliqués dans les applications agronomiques. De plus, afin d’assurer à la fois l’utilisation sûre et l’acceptation de cette nouvelle opportunité, les effets indésirables sont également discutés.
Mots clés:
stress abiotique ; rendement des cultures ; champ magnétique ; eau magnétisée ; magnéto-amorçage ; germination des graines
1. Introduction
La germination des graines est améliorée par des traitements de pré-semis par des méthodes chimiques ou physiques en brisant la dormance, ce qui protège les graines contre les ravageurs et les maladies et permet un établissement uniforme des cultures sur le terrain. La vigueur et la vitalité des semences sont perdues lors du stockage en raison de la détérioration, ce qui entraîne finalement la perte de matériel de semence coûteux. Les traitements d’amorçage tels que l’osmo-amorçage, l’hydro-amorçage, l’amorçage halo et l’amorçage à matrice solide sont des techniques de pré-semis qui hydratent les graines pendant le processus de traitement, améliorant ainsi la germination au détriment du stockage des graines [ 1 , 2]. Ces graines ne peuvent pas être stockées plus longtemps et doivent être semées immédiatement après le traitement. En gardant cet inconvénient à l’esprit, l‘amorçage de la graine avec un champ magnétique (MF) peut constituer une alternative viable pour améliorer la vigueur des graines [ 1 ].
Au cours des dernières décennies, l’environnement humain a été modifié en raison de l’utilisation extrême du MF. L’utilisation excessive et constante d’appareils électroménagers, d’instruments médicaux, de véhicules de transport et d’équipements de communication expose les plantes (plus que toute autre espèce) à une plus grande quantité de MF [ 3 ]. En effet, l’intensité du champ géomagnétique qui affecte la croissance des plantes par MF naturel avec une intensité de 50 à 60 μT [ 4] augmente et les progrès technologiques modifient cette intensité MF. Par conséquent, lorsqu’elles sont exposées au MF naturel, les plantes réagissent différemment en fonction de l’intensité du MF, ce qui peut influencer positivement ou négativement leur développement [ 5 ]. Les effets du MF dans les semences et les plantes ont fait des progrès impressionnants au cours des 10 dernières années en phytologie. Actuellement, il existe des preuves réelles que le traitement magnétique de pré-germination des graines avant le semis permet de réduire les coûts de plantation car les taux de germination sont considérablement augmentés, tout en favorisant la croissance des plantes [ 6 , 7 , 8 ] . Cependant, d‘autres ont montré que le développement est inhibé [ 9 , 10 ].
Diverses pratiques agronomiques ont souvent été employées pour améliorer le prétraitement de germination des graines, qui stimule la germination, la levée et la vigueur des graines. Cet article met en avant le MF comme une alternative aux traitements conventionnels à base de substances chimiques (régulateurs de croissance des plantes), renforçant l’importance de comprendre les différentes interactions entre les champs électromagnétiques et les processus physiologiques des plantes [ 11 , 12 , 13]. Les rendements des cultures en général et l’homogénéité sont augmentés avec l’application de substances chimiques (telles que les hormones) dans le traitement des semences avant le semis, bien qu’elles soient considérées comme très efficaces et invasives, écologiquement incorrectes et difficiles à appliquer. Au cours des dernières décennies, la découverte d’hormones et d’engrais chimiques a joué un important facteur d’amélioration du rendement dans la culture des plantes, mais l’utilisation actuelle d’engrais chimiques et d’autres matériaux a été controversée, ce qui a orienté l’utilisation de traitements alternatifs respectueux de l’environnement avec un coût réduit. , telles que les rayons gamma, le laser, le faisceau d’électrons, les micro-ondes, les MF et les radiofréquences pour provoquer la bio-stimulation des graines [ 14 , 15]. Ainsi, le traitement biophysique peut être considéré comme une alternative car il réduit la quantité de toxine dans les plantes ou les produits de plantes et entraîne une augmentation de la sécurité alimentaire et environnementale [ 14 ]. Le traitement des semences par champ magnétique est devenu très populaire dans le secteur agricole. Le traitement des semences avant le semis avec MF, appelé « magnétopriming », est un traitement de pré-semis non destructif et sec qui a été signalé comme augmentant le taux de germination et la vigueur des semis de nombreuses cultures [16 , 17 , 18 , 19 ] . Il existe plusieurs rapports sur les changements métaboliques survenant lors de la germination des graines en réponse à la magnétoprimation dans des environnements non stressés [ 16 , 17 ,18 , 19 ]. Les effets de la bio-stimulation magnétique des graines sous stress salin à l’aide de MF stationnaire ont été rapportés par Thomas et al. [ 20 ] et Kataria et al. [ 21 ].

Un autre avantage de l’utilisation de MF est lié à l’augmentation des taux de germination et à l’augmentation possible de la perméabilité membranaire, facilitant le processus d’absorption d’eau par les graines [ 22 ]. Il existe encore d’autres études qui ont confirmé l’effet de l’eau magnétisée dans les plantes ou les graines. En ce sens, Maheshwari et Grewal [ 23 ] et Hilal et al. [ 24 ] ont déclaré que MF peut affecter l’absorption d’eau et de nutriments, ainsi que l’amélioration de la croissance des plantes. Au-delà de l’amélioration des taux de germination, les graines exposées au MF induisent des effets positifs tels qu’une capacité de prolifération cellulaire accrue, ce qui induit éventuellement une croissance rapide des plantes [ 25]. D’autres chercheurs ont également signalé l’effet positif du MF sur l’augmentation de la germination des graines, la vigueur des semis, les pigments photosynthétiques, l’efficacité du photosystème II (PSII) et l’indice de performance basé sur l’absorption de l’énergie lumineuse, ainsi que sur la promotion d’une photosynthèse efficace et l’atténuation les effets néfastes du stress salin, hydrique et UV-B dans le soja ( Glycine max L. Merr. Var: JS-335) [ 21 , 26 , 27 , 28 ].
Dans certaines études, le MF pourrait réduire les dommages oxydatifs des plantes, en raison des activités d’enzymes antioxydantes telles que la peroxydase, la polyphénol oxydase (PPO), la superoxyde dismutase (SOD) et la catalase (CAT) dans les cellules végétales. Plus précisément, MF a affecté l’activité antioxydante et augmenté l’activité des ions radicaux libres dans les plantes [ 29 , 30 , 31 ]. D’autres études ont rapporté de multiples effets négatifs du MF sur les plantes. Ces effets comprennent l’inhibition de la croissance cellulaire, l’augmentation des radicaux libres, l’augmentation de la lignine et de la subérine sur les parois des cellules, et la réduction de la germination des graines et de la croissance des organes [ 32]. De plus, le MF s’avère être un outil potentiel en agriculture lorsqu’il est utilisé comme pesticide. Par exemple, Mahajan et Pandey [ 6] ont conclu que l’utilisation alternative du MF à l’utilisation d’insecticides et de pesticides est possible, dans laquelle ils ont utilisé la technique de magnéto-amorçage dans les semences pour prévenir les ravageurs et les maladies agricoles, obtenant un rendement élevé de haricots mungo après la plantation. Les résultats étaient comparables à l’application d’engrais chimiques, d’insecticides et de pesticides afin de protéger les plantes du virus de la mosaïque jaune du haricot mungo. Les chercheurs ont alors supposé que le magnéto-amorçage était prometteur, s’imposant comme une technique efficace, propre et abordable, induisant à la fois une résistance des plantes et une productivité élevée. La présente revue se concentre sur l’application de MF et MW (eau magnétisée) dans la germination des graines, la croissance des plantes et le développement des plantes et des microalgues.
2. Les effets de l’application MF sur le développement des plantes
2.1. Effets des traitements magnétiques sur la germination des graines
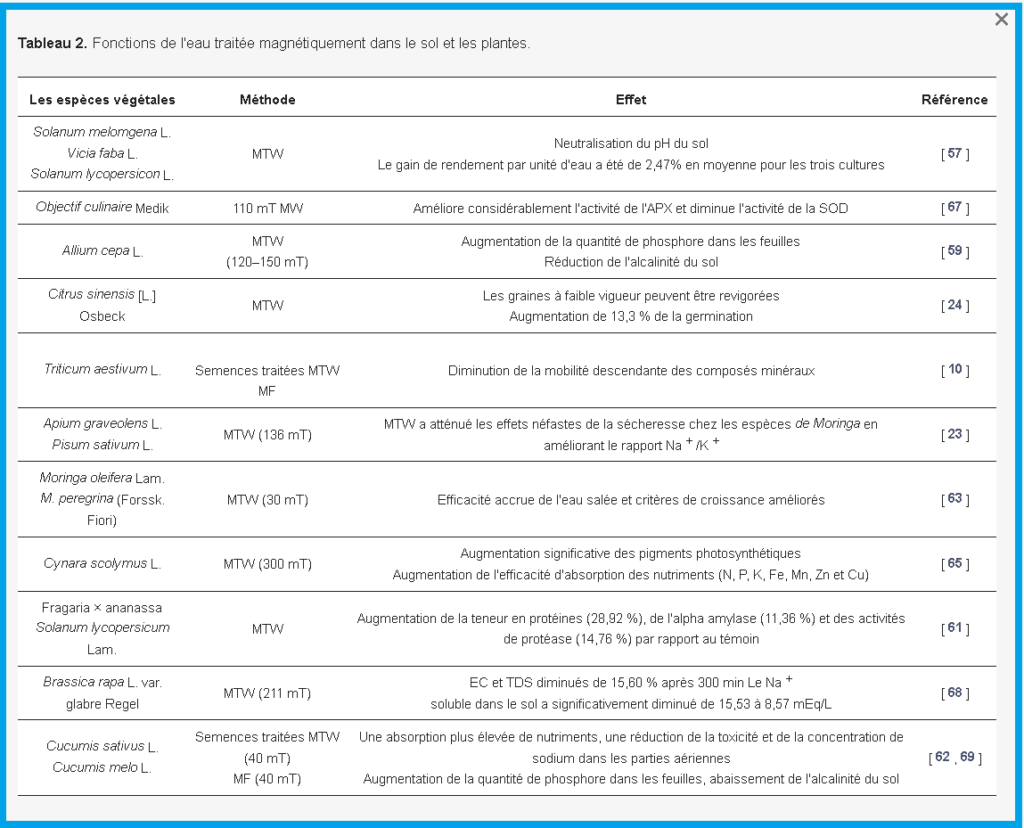
Les plantes qui poussent sur la terre sont affectées par le MF naturel avec une intensité de 50 à 60 µT [ 33 ]. L’effort pionnier pour améliorer le rendement en graines en l’exposant aux MF et aux champs électromagnétiques (EMF) a été réalisé depuis 1930 [ 34 ]. L’effet de l’exposition à un MF sur la germination des graines a été l’objectif de nombreuses études, et l’amélioration de la germination des graines due à l’exposition au MF a été confirmée par de nombreux scientifiques [34 , 35 ] .
Mahajan et Pandey [ 6 ] ont évalué les impacts des champs magnétiques statiques (SMF) sur la germination du haricot mungo ( Vigna radiata(Linn.) Wilczek.) et ont signalé une augmentation linéaire du taux de germination moyen, du coefficient de taux de germination et de l’absorption d’eau avec l’augmentation de l’intensité MF.
De plus, Menegatti et al. [ 8 ] ont indiqué que l’exposition des graines de fruit de la passion au MF de manière isolée stimulait la germination, l’émergence et la vigueur des graines.
Les graines de pois chiches traitées magnétiquement ( Cicer arietinum L.) ont montré une amélioration des performances des graines en termes de vitesse et de longueur de germination et de poids sec des semis, et la réponse variait en fonction de l’intensité du champ et de la durée d’exposition [ 36 ].
Les mêmes effets positifs sur le taux de germination des graines et l’indice de vigueur ont été trouvés dans les graines de concombre ( Cucumis sativus L. Var. Barsati), de laitue ( Lactuca sativaL.), graines de maïs ( Zea mays L. Var. HQPM-1), graines de tomate ( Solanum lycopersicum L. Var. MST/32) et graines de radis ( Raphanus sativus L.) par certains chercheurs [ 17 , 22 , 37 , 38 , 39 ].

De plus, des études ont également confirmé que les graines traitées magnétiquement poussent plus haut et plus lourdes que le témoin, et augmentent même l’absorption d’eau [ 22 ]. Ils ont également des racines plus profondes et plus vigoureuses que le témoin [ 14 , 16 , 19 ].
De plus, Florez et al. [ 40] ont observé une augmentation du taux d’allongement des semis de blé traités par MF.
D’autre part, Belyavskaya [ 12 ] a signalé une augmentation de l’intensité et des émissions de dioxyde de carbone (CO 2 ) de 70 à 100 % lorsque les semis d’orge ( Hordeum vulgare L.) étaient traités par 10 µT MF.
De plus, Kavi [ 41 ] a rapporté qu’une application appropriée de MF réduit le potentiel d’hydrogène dans la paroi cellulaire, prévient la dormance des graines, influence le métabolisme des cellules méristématiques, augmente l’absorption des nutriments et améliore la capacité photosynthétique [ 34 , 41 ]. Avec tout cela, les mécanismes exacts dans lesquels MF affecte la germination des graines sont encore inconnus, avec seulement quelques publications sur cet aspect.
Les scientifiques ont découvert que le MF améliore la germination des graines en modifiant les processus biochimiques en stimulant l’activité des protéines et des enzymes [ 30 , 42 ]. En fait, MF peut interagir avec le champ électrique interne des systèmes biologiques grâce à son comportement de résonance. Les cellules vivantes possèdent des charges électriques exercées par des ions ou des radicaux libres, qui agissent comme des aimants endogènes et ont été impliqués dans les processus biochimiques [ 43 ]. Ainsi, le traitement MF externe augmente l’absorption d’ions et améliore donc la valeur nutritionnelle [ 43]. D’autres études ont indiqué que le MF interagit avec le courant ionique dans la membrane de la cellule embryonnaire. Cette interaction modifie la concentration en ions et la pression osmotique fournies des deux côtés de la membrane, modifiant ainsi la relation entre l’eau et les graines [ 22 ].
De plus, des recherches récentes d’Anand et al. [ 19 ] ont montré que les plantes répondaient à différents MF en modifiant leur expression génique et leur phénotype. En fait, ils ont rapporté que RACK1 (c’est-à-dire le récepteur de la protéine kinase activée C1) et la métallothionéine jouent un rôle important dans la voie de transduction du signal médiée par les espèces réactives de l’oxygène (ROS) pour améliorer la vitesse de germination dans les graines de tomates magnétoprimées ( Lycopersicon esculentum L . Mill. var. Pusa Rohini). Racuciu et al. [ 44 ] ont rapporté que dans une étude menée sur des racines poilues induites par Agrobacteriumspp. (pas de vraies racines ni de méristème racinaire), l’exposition à 50 µT MF a eu un effet stimulant sur l’augmentation du poids frais, de la teneur en pigments photosynthétiques et de la teneur en acides nucléiques, et a augmenté la longueur des semis de maïs ( Zea mays L. ) . Cependant, le traitement MF avec une induction plus élevée (100 à 250 µT) a eu un effet inhibiteur sur les paramètres susmentionnés. L’ intensité MF et son effet sur plusieurs espèces de plantes sont résumés dans le tableau 1 .
D’autres études ont montré que le SMF provoque l’apoptose induite dans les cellules de tabac cultivé en suspension ( Nicotiana tabacum L. cv. Burley 21), et la réduction de la croissance du basilic ( Ocimum basilicum L.) [ 9 , 51 ]. De plus, Kordas [ 5 ] a signalé que la MF provoquait une légère diminution de la longueur de la tige du blé, tandis que le rendement en grain et la quantité de paille étaient légèrement augmentés. Contrairement aux études susmentionnées, certains ne sont pas d’accord avec la MF [ 12 , 52 ]. Par exemple, Ijaz et al. [ 10] ont trouvé que les graines de blé cv. NR-234 avec une faible viabilité (45%), lorsqu’il est soumis à des traitements de magnétisation, n’a montré aucune augmentation de la germination, et que ce traitement était insignifiant. De plus, il a été observé qu’un MF faible allant de 100 à 0,5 µT affecte négativement la germination des graines, la croissance des semis des plantes, des racines et la division cellulaire dans le méristème racinaire [ 53 ] .
2.2. Effets du MW sur la germination des graines et la croissance des plantes
Il est bien connu que des approvisionnements en eau adéquats, ainsi que la capacité d’absorption d’eau des plantes et la qualité des graines, sont considérés comme les facteurs les plus importants pour la croissance d’une plante. Ceci est particulièrement important en ce qui concerne la demande mondiale de plus de nourriture à partir de ressources en eau moindres [ 54 ]. Par conséquent, une approche scientifique est nécessaire pour maintenir la productivité des cultures agricoles.
Le traitement de l’eau par MF est un autre aspect particulier de l’utilisation des MF et est l’une des techniques physiques prometteuses pour améliorer la qualité de l’eau et la productivité des cultures.
L’eau traitée magnétiquement peut améliorer la production agricole, ainsi que la germination des graines [ 55 ], accélérant la croissance végétative des semis, ce qui améliore également la teneur en minéraux des graines et des fruits [ 54]. L’énergie magnétique pourrait améliorer les propriétés physiques et/ou chimiques de la qualité du sol et de l’eau.
L’exposition de l’eau à un champ magnétique entraîne des altérations de ses propriétés de base telles que la force ionique, le pH et la force de tension superficielle, offrant une plus grande capacité de mouvement en intensifiant la vibration interne des molécules d’eau, ce qui améliore l’effet polarisant, entraînant une augmentation de l’absorption d’eau dans la cellule [ 56 ]. Kareem [ 57 ] a évalué l’effet de l’irrigation à l’eau magnétisée sur le pH du sol. Leurs résultats ont montré que le pH diminuait jusqu’à des niveaux proches de la neutralité. Cette méthode peut réduire la période de croissance des cultures et économiser plus d’eau d’irrigation.
Hirota et al. [ 58 ] ont montré que lorsque les graines de concombre cultivées étaient irriguées par de l’eau magnétisée, il en résultait une augmentation de la croissance de ces plantes par rapport aux plantes témoins.
Dans une autre étude, Fernandez et al. [ 59 ] ont rapporté que les semis élevés par MW étaient plus forts et plus sains en augmentant l’absorption des nutriments de l’eau des plantes.
La productivité de l’eau a été augmentée de 1,65, 1,70 et 1,88 pour l’aubergine ( Solanum melomgena L. cv. Florida High Bush), la tomate ( Solanum lycopersicon L. cv. Logaen) et la féverole ( Vicia fabaL.cv. Isban), respectivement, par l’eau traitée magnétiquement (MTW). Cela a conduit à des économies d’eau de 11 %, 14,2 % et 13,5 % pour les trois cultures, respectivement, augmentant finalement le bénéfice net.
En utilisant la technique de traitement magnétique, les rapports entre le rendement net par unité d’eau et celui de l’utilisation d’eau non traitée étaient de 1,97, 2,45 et 3,0 pour l’aubergine, la tomate et la fève, respectivement [ 57 ] .

D’autres études ont confirmé les effets de l’eau magnétisée et ont indiqué que la quantité de phosphore dans les feuilles d’agrumes augmentait lorsqu’elles étaient traitées par MW [ 24 , 60 , 61 ]. Cela peut affecter l’absorption du calcium (Ca) et du phosphore (P) dans les agrumes, et comme les plantes peuvent y accéder plus facilement, la croissance de la plante sera améliorée [ 23 ].
En tant que tel, les chercheurs ont conclu que MTW modifie les relations hydriques dans le grain, et cet effet peut expliquer en partie l’accélération du métabolisme des graines et du taux de germination [ 12 , 14 ]. Selon un rapport d’Ijaz et al. [ 10 ], graines de blé ( Triticum aestivumL.cv. NR-234) avec une faible viabilité (45%) lorsqu’ils sont soumis à des traitements MW ont été revigorés. La stabilité du pH de l’eau absorbée par les graines ou les plantes peut avoir été atteinte en raison de l’altération de son ionicité, résultant de la rupture des liaisons hydrogène présentes dans la molécule, ce qui a permis à l’eau d’être présente à une concentration plus élevée d’ions libres.
En comparant les composants des sols irrigués par de l’eau magnétisée et des sols irrigués par l’eau du robinet, Noran et al. [ 62 ] ont observé une différence de concentration en Ca, P, azote (N), potassium (K), sodium (Na) et magnésium (Mg). Ils ont déclaré que MW diminue la mobilité vers le bas des composés minéraux, ce qui est dû au processus de cristallisation accélérée et au dépôt des éléments minéraux [ 54 ]. Maheshwari et Grewal [23 ] ont rapporté les effets de MW dans la réduction du pH dans le sol, ce qui a entraîné une absorption plus élevée de nutriments. Là où il y a eu une augmentation des concentrations de Ca et de P dans le pois ( Pisum sativum L.) et le céleri ( Apium graveolens L.), il y a eu une restriction de charge de Na, ainsi qu’une réduction de la toxicité et de la concentration de Na dans les parties aériennes. .
De plus, la technique magnétique nous permet d’utiliser efficacement l’eau salée (teneur en sel de 2000 ppm et jusqu’à 5000 ppm) pour l’irrigation des cultures [ 54 ]. Par conséquent, Hasan et al. [ 63 ] ont rapporté que l’utilisation de MW chez deux espèces de Moringa ( Moringa oleifera Lam. et Moringa peregrina(Forssk.) Fiori), cultivée sous stress hydrique salin, a permis la récupération de l’inhibition de la croissance induite par la restriction hydrique, la chlorose et la perturbation ionique. De plus, certains chercheurs ont révélé une réponse notable au stress lors de l’irrigation de Silybum marianum (L.) Gaertn. plantes par de l’eau de mer traitée magnétiquement [ 64 ].
Le traitement MF réduira les effets nocifs du stress de salinité au stade précoce des semis et réduira les dommages oxydatifs, conduisant à des améliorations des attributs physiologiques pour la croissance des plantes sous stress d’irrigation par l’eau de mer [ 64 ] . Bagherifard et Ghasemnezhad [ 65 ] ont déclaré que l’exposition à l’eau salée améliorait différents critères de croissance de l’artichaut ( Cynara scolymusL.) feuilles.
D’autres chercheurs ont observé qu’une augmentation du taux de germination des graines de laitue ( Lactuca sativa L.) traitées avec MF statique (0–10 mT) était cohérente avec le taux d’eau absorbée des graines [ 66 ]. La méthode de traitement magnétique de l’ eau dans le sol et les plantes et ses effets sur plusieurs plantes sont résumés dans le tableau 2 .
2.3. Effets des MF sur la réduction des dommages oxydatifs
De nombreux chercheurs ont révélé les effets du MF sur les activités des enzymes antioxydantes telles que la peroxydase (POD), la polyphénol oxydase (PPO), la superoxyde dismutase (SOD) et la catalase (CAT) dans les cellules végétales. À cet égard, Bhardwaj et al. [ 17 ] ont observé une augmentation des activités des enzymes antioxydantes, à savoir la SOD de 8 %, la CAT de 83 % et la glutathion réductase (GR) de 77 % dans les graines de concombre ( Cucumis sativus L.) exposées à un SMF, par rapport à au contrôle. Une expérience similaire réalisée sur plusieurs espèces végétales, dont l’artichaut ( Cynara scolymus L.) et Zea mays L. (génotype = single cross 704), a rapporté que l’exposition au MF provoquait une augmentation significative des activités de la CAT et de l’ascorbate peroxydase (APX) [65 , 70 ]. Hajnorouzi et al. [ 71 ] ont conclu que le prétraitement par MF alternatif favorisait la croissance des semis de maïs en atténuant la production excessive d’espèces réactives de l’oxygène. L’activité de la SOD dans les semis traités magnétiquement a diminué, tandis que la capacité antioxydante totale de ces semis a augmenté par rapport au témoin.
Il a été révélé que, comme les cellules humaines et animales, les cellules végétales peuvent être influencées par la MF. De plus, les effets des MF ont été liés au découplage du processus radicalaire dans les membranes [ 29 , 30 , 31 ]. De plus, les lipides des membranes cellulaires sont sujets aux dommages oxydatifs car certains radicaux libres ont tendance à se concentrer sur la membrane et à provoquer des dommages oxydatifs connus sous le nom de « peroxydation lipidique » [72 , 73 , 74 , 75 ] . L’accumulation de ces radicaux libres peut provoquer un stress oxydatif [ 32 , 53 , 76], et le stress oxydatif provoque une modification de l’activité enzymatique, de l’expression des gènes et de la libération de calcium des réserves intracellulaires. De plus, ce stress peut affecter la structure membranaire et la croissance cellulaire, et induire la mort cellulaire (apoptose) [ 71 , 77 , 78 ]. En traitant les graines de soja par les SMF, Shine et Guruprasad [ 79 ] ont observé une augmentation de la production de ROS médiée par la peroxydase de la paroi cellulaire, tandis que la production de teneur en acide ascorbique et les activités SOD et APX étaient diminuées dans la partie hypocotyle de la germination. graines.
De nombreuses expériences réalisées sur plusieurs espèces végétales, dont le soja ( Glycine max L. Merr. Var : JS-335), le maïs ( Z. maysL.Var. HQPM-1) et la tomate ( L. esculentum ) ont rapporté que le prétraitement avec MF induit une plus grande résistance des plantes plus tard dans des conditions de stress hydrique, suggérant que cette efficacité est liée à l’amélioration du système antioxydant [ 26 , 37 , 80 ]. Shabrangi et Majd [ 81 ], enquêtant sur les effets du stress hydrique chez la lentille ( Lens culinarisMedik.), ont observé qu’il y avait plus de résistance chez les semis prétraités par des champs magnétiques, avec une augmentation significative des activités APX et SOD dans les racines et les pousses. Ces enzymes antioxydantes récupèrent les ROS et autres modifications chimiques produites dans les cellules soumises à un stress. Les résultats suggèrent que les semences prétraitées par les MF leur permettent de surmonter les facteurs environnementaux nocifs. Çelik et al. [ 82] ont étudié deux enzymes du système de défense, les activités SOD et CAT sous application MF. Les résultats ont indiqué que la fonction des enzymes de défense dans les semis était intensifiée en raison du traitement par le MF, ce qui indiquait que pour les cellules végétales, un MF crée une condition de stress similaire à d’autres facteurs de stress environnementaux. Cependant, les augmentations des temps d’exposition au MF ne provoquent pas d’augmentations linéaires des activités enzymatiques dans les études in vitro et in vivo.
De même, certaines études ont rapporté des effets négatifs du MF sur les plantes. Roux et al. [ 83 ] ont exposé des plants de tomates à des MF pendant une courte période (10 min), et quelques minutes après la stimulation électromagnétique, l’ARNm lié au stress (calmoduline, protéine kinase dépendante du calcium et inhibiteur de protéinase) s’est accumulé rapidement, et 30 min après la stimulation électromagnétique. traitement, la concentration d’ATP et la charge énergétique de l’adénylate ont diminué de manière transitoire. Cela suggère fortement qu’ils sont le résultat direct de l’application de MF et que ce rayonnement est considéré comme un stimulus nocif par les plantes. Ces résultats contradictoires de ces études peuvent dépendre des caractéristiques de l’exposition au champ, telles que l’intensité et la durée.
2.4. Atténuation des stress abiotiques
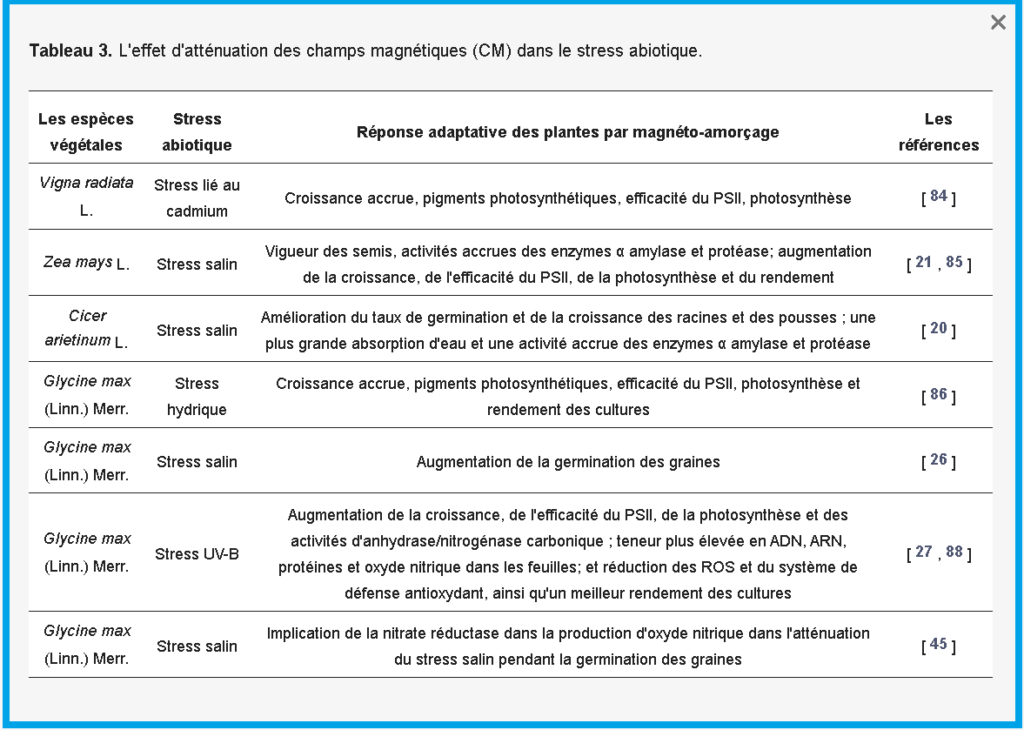
Bien que les stress abiotiques, tels que la salinité, les UV-B et le stress hydrique, réduisent la vigueur des semis, le taux de germination, la nodulation, le taux de croissance de la biomasse, le métabolisme du carbone et de l’azote, qui diminuent tous le rendement des cultures, il a été prouvé que les plantes présentent des performances compensatoires sur tous ces paramètres contre les stress abiotiques ainsi que dans des conditions sans stress [ 1 ].
El-Yazied et al. [ 80] ont obtenu une augmentation significative du pourcentage de germination et une réduction du temps nécessaire à la germination de la tomate. Ils ont également observé, chez les semis issus de semences traitées, une augmentation de la longueur de la tige, du diamètre de la tige, de la surface foliaire et du poids frais et sec, même dans des conditions salines.
Plusieurs études ont montré que le SMF peut augmenter la germination des graines et la vigueur des semis sous stress salin et de métaux lourds dans le pois chiche, le soja, l’orge, le haricot mungo et le maïs [20 , 21 , 47 , 84 , 85 ] .
Des résultats similaires obtenus par Baghel et al. [ 26 ] ont démontré l’efficacité de la magnéto-amorçage, des résultats positifs sur les attributs de croissance des plantes, le nombre de nodules racinaires, les nodules, le poids frais et l’accumulation de biomasse dans le soja ( Glycine max L. Merr. Var : JS-335), soit exposé ou non exposé au stress salin. Les résultats suggèrent en outre que l‘utilisation de MF a augmenté le métabolisme du carbone et de l’azote et amélioré le rendement du soja en termes de nombre de gousses, de nombre de graines et de poids des graines dans des conditions salines et non salines. Baghel et al. [ 86], évaluant l’effet du SMF sur les réponses morphologiques et physiologiques du soja au stress hydrique, a conclu que le prétraitement des graines par MF entraîne une augmentation des pigments photosynthétiques, de l’efficacité du PSII et de l’indice de performance basé sur l’absorption de l’énergie lumineuse, et favorise l’efficacité de la photosynthèse et atténué les effets néfastes du stress hydrique sur le soja. Un prétraitement supplémentaire au SMF a amélioré l’efficacité du PSII, le taux de photosynthèse et le rendement des cultures sous des stress abiotiques tels que les UV-B et le stress salin en diminuant les ROS [27 , 28 , 87 ] . Anand et al. [ 87] ont expliqué l’atténuation des effets néfastes du stress hydrique par le fait que le MF réduisait les productions de radicaux libres et l’activité enzymatique antioxydante. La réponse adaptative des plantes par magnétoamorçage sous stress abiotique chez plusieurs espèces végétales est résumée dans le tableau 3 .
Le prétraitement avec SMF dans le soja exposé au stress salin a également eu un effet positif sur l’augmentation des activités α-amylase, protéase et NR, ainsi que des niveaux plus élevés de H 2 O 2 , O 2 •− et d’oxyde nitrique (NO) [ 45 ]. Les auteurs ont suggéré le NO comme l’une des principales molécules de signalisation dans la tolérance au sel induite par MF chez les semis de soja. Nous avons essayé de clarifier comment le champ magnétique atténue les stress abiotiques chez les plantes ( Figure 1 ).
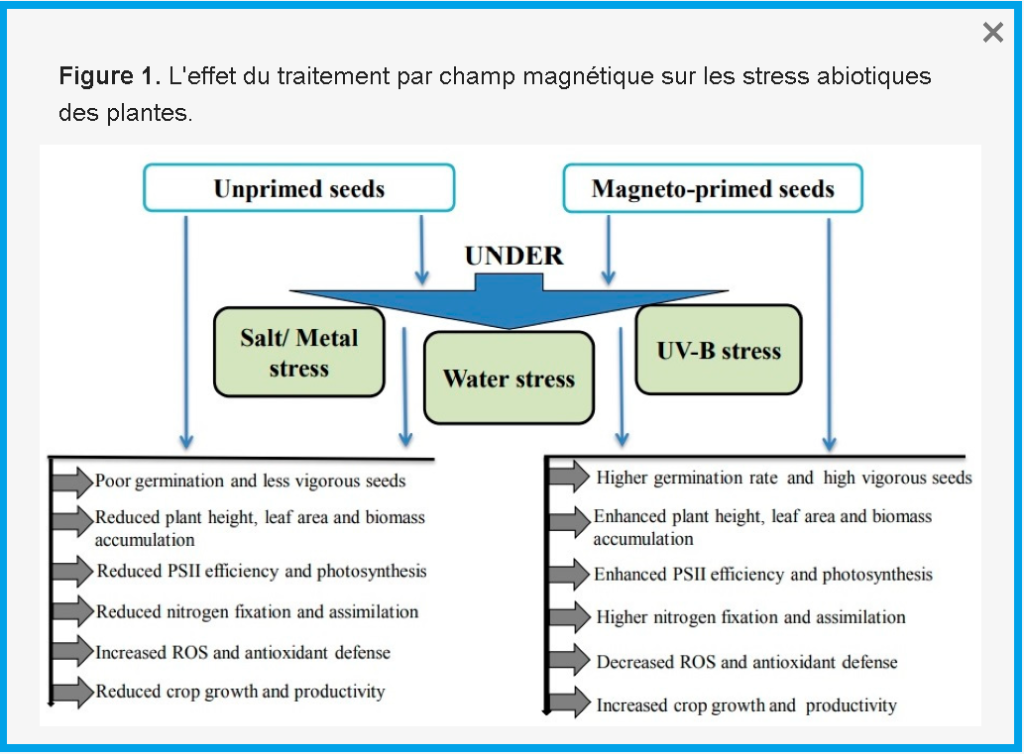
3. Les effets des MF sur les microalgues
Les microalgues sont des micro-organismes qui peuvent être procaryotes ou eucaryotes, tout comme les plantes sont des êtres vivants photosynthétiques. Plusieurs études ont vérifié l’influence de l’application de MF pendant les cultures et des résultats positifs ou négatifs sur la croissance ou la production de composés ont été observés [ 89 ].
Chez les microalgues, ces effets dépendent de l’état physiologique de la cellule, du type de cellule (procaryote ou eucaryote), du temps d’exposition, de l’intensité, de la forme d’application et du type de dispositif générateur de champ. Certains auteurs rapportent des explications possibles sur la façon dont le MF affecte la croissance des microalgues, et depuis 2010, le nombre d’études a augmenté.
Petit et al. [ 90] ont cité que le MF modifie les concentrations de radicaux libres en raison d’un stress oxydatif accru, ce qui est le mécanisme le plus probable de l’effet du MF sur les microalgues. Beruto et al. [ 91 ] ont conclu que les champs électromagnétiques à basse fréquence (EM-ELF) n’agissaient pas sur la division mitotique, mais jouaient un rôle important dans la promotion de la clusterisation cellulaire en phase liquide. Wang et al. [ 92 ] ont observé que le MF augmente la croissance des microalgues et régule son système de défense antioxydant pour protéger efficacement les cellules. Tu et al. [ 93 ] ont vérifié que la MF stimule la croissance des algues et la production d’oxygène en utilisant Scenedesmus obliquus cultivé dans les eaux usées municipales. Luna et al. [ 94] ont observé un changement de modification dans la disposition des tilacoïdes chez la cyanobactérie Synechocystis aquatilis induite par MF (37,7–44,3 mT), faisant varier la distribution et les distances entre les tilacoïdes et les membranes tilacoïdes. D’autres explications possibles ont été rapportées par Santos et al. [ 89 ]. Les études se sont concentrées sur la culture de différentes microalgues avec l’application de MF et ont vérifié quels sont les effets observés. Les microalgues déjà évaluées sont Nannochloropsis oculata [ 95 ], Chlorella fusca [ 96 , 97 ], Spirulina sp. [ 98 , 99], et d’autres espèces de microalgues citées dans le tableau 4 .
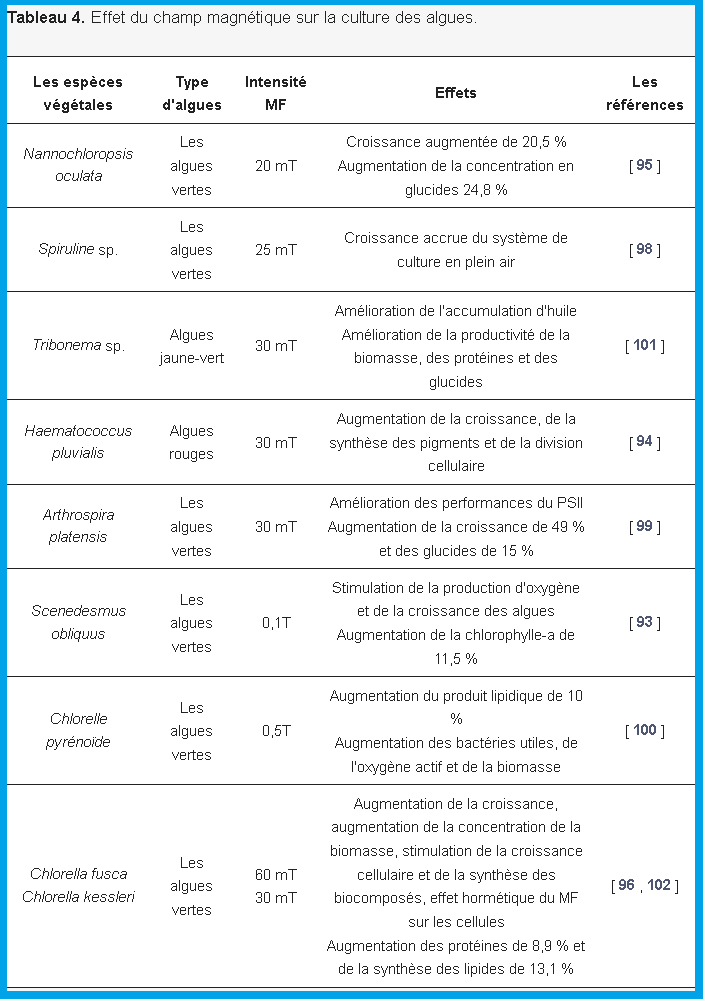
Les effets observés sont différents selon les conditions de l’application MF.
Luna et al. [ 94 ] ont observé qu’avec Chlorella vulgaris , l’application continue de MF favorise l’accumulation de glucides, de lipides et de protéines dans la phase de croissance exponentielle et de lipides dans la phase stationnaire. Deamici et al. [ 96 ] ont vérifié que 60 mT appliqués tout au long de la culture augmentaient respectivement de 20,5 % et 24,8 % en termes de biomasse et de teneur en glucides. Han et al. [ 100 ] ont observé que 0,5 T stimulait 12 % de biomasse et 10 % de productivité lipidique lorsque la MF était appliquée tout au long de la culture. Chu et al. [ 95 ] ont vérifié les effets de la concentration en MF et en nitrate sur la croissance desNannochloropsis oculé .
Lors de l’utilisation de 20 mT et 150 mg L −1 de nitrate, le taux de croissance spécifique maximal et la productivité lipidique maximale ont augmenté de 166 % et 103 %, respectivement. Huo et al. [ 101 ] ont observé que 30 mT peuvent affecter la composition biochimique de Tribonema sp. et améliorer l’accumulation d’huile.
4. Mécanismes possibles de magnéto-amorçage
Dans les systèmes biologiques, un certain nombre d’hypothèses ont été proposées pour les mécanismes de perception et de réponses MF, parmi lesquelles le modèle de résonance cyclotronique ionique, le modèle de résonance paramétrique, les excitations quantiques cohérentes et les mécanismes de spin et de radicaux libres recevant actuellement plus d’attention [ 12]. Les réactions catalysées par des enzymes impliquant des intermédiaires avec des paires de radicaux libres sont également appelées « mécanisme de paire radicalaire » (RPM). La modulation du taux de conversion singulet-triplet des paires de radicaux libres est affectée par un champ magnétique faible. Dans le processus de la chaîne respiratoire mitochondriale, les porphyrines produisent des radicaux libres, qui peuvent également être influencés par un champ magnétique externe car il affecte les conversions singulet-triplet. Le RPM est actuellement le seul mécanisme physiquement plausible indiquant le rôle du cryptochrome en tant que candidat à la réception magnéto qui se traduit par la génération de paires de radicaux flavine-trytophan [ 35]. Les cryptochromes sont des récepteurs de protéines de saveur légère et on pense qu’ils sont impliqués dans la magnétoperception des plantes car ils peuvent former des paires radicales de flavine adénine dinucléotide réduite (FADH2) et de résidu de tryptophane dans la structure protéique après exposition à la lumière bleue [ 103 , 104 ] .
Sur la base de l’hypothèse du ferrimagnétisme, la perception du SMF par les plantes est obtenue par son effet direct sur les particules de fer, les composés de fer minéraux (par exemple, Fe 3 O 4 et Fe 3 S 4 ) et les protéines contenant du fer [ 104 , 105 ] . La phytoferritine est présente dans les cellules végétales sous forme de magnétite cristalline (Fe 3 O 4 ), ε-Fe2 O 3 , et l’hématite (α-Fe 2 O 3 ) [ 106 ], et peuvent interagir plus fortement avec les champs magnétiques qu’avec les matériaux diamagnétiques ou paramagnétiques. Ces particules peuvent également affecter les radicaux libres générés par les superoxydes [ 107 ].
Plusieurs rapports indiquent que la MF peut provoquer une surproduction de ROS et l’initiation d’un stress oxydatif. De plus, l’influence du SMF a également altéré les activités des antioxydants enzymatiques ou l’expression de leurs gènes [ 104 , 108 , 109 , 110]. L’autre théorie de la « résonance cyclotronique des ions » tourne autour du fait que les ions doivent circuler dans un plan perpendiculaire à un champ magnétique externe avec leurs fréquences de Lamor, qui peuvent interférer avec un champ électromagnétique alternatif [ 34 , 111 ] .
Les effets biologiques peuvent être élucidés comme une interaction entre le MF et le courant ionique dans la membrane cellulaire de l’embryon végétal, ce qui provoque des altérations des concentrations ioniques et de la pression osmotique des deux côtés de la membrane [112 ] . Les changements dans les mécanismes d’absorption d’eau sont causés par des altérations des flux ioniques à travers la membrane cellulaire [ 66]. Les imbibitions de graines traitées magnétiquement ont montré une hydratation plus rapide des macromolécules et des membranes et une plus grande activité d’enzymes telles que l’α-amylase et la nitrate réductase pendant la germination des graines, qui sont responsables d’une germination plus rapide des graines magnéto-amorcées par rapport aux graines non exposées [ 45 , 48 , 113 , 114 , 115 , 116 , 117 ]. Les divers effets morphologiques, physiologiques et biochimiques du prétraitement des semences par champ magnétique sur les plantes sont représentés à la figure 2 .
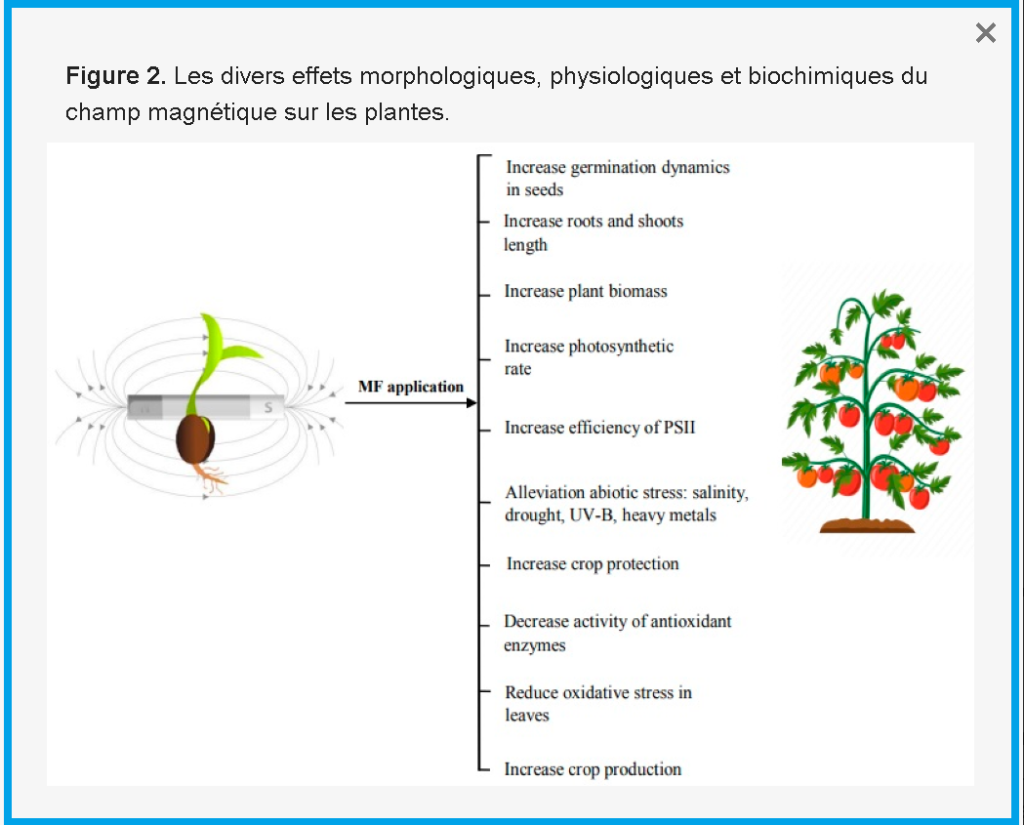
5. Conclusions et perspectives
Pour augmenter la germination des graines, la productivité et le développement des cultures, une bonne combinaison d’intensité MF et de temps d’exposition est essentielle.
De nombreuses études ont prouvé que ses impacts positifs peuvent améliorer la germination des graines, la longueur des racines et des pousses, l’absorption de l’eau et du CO 2, la teneur en pigments photosynthétiques, et enfin l’augmentation de la production agricole même sous stress abiotiques.
D’autre part, d’autres chercheurs ont montré que les paramètres susmentionnés n’ont pas été améliorés, mais ont plutôt diminué depuis, et le MF provoque une inhibition de la croissance des plantes. Le mécanisme par lequel les plantes perçoivent les MF et régulent la voie de transduction du signal n’est pas entièrement compris. Il a été suggéré que la perception/signalisation MF chez les plantes est médiée par les photorécepteurs de lumière bleue, les cryptochromes. Il a également été découvert que ROS et NO sont les molécules de signalisation pour la germination des graines induite par magnétoamorçage.
Cependant, cet aspect de la magnétobiologie mérite encore une enquête approfondie, ainsi que les effets secondaires génotoxiques potentiels des MF. Tous ces travaux ont mis en évidence la nécessité d’études supplémentaires pour étendre nos connaissances sur les mécanismes moléculaires impliqués dans la fixation de la germination des graines, l’augmentation de la vigueur des semis et l’amélioration de la capacité photosynthétique des plantes magnéto-amorcées sous stress abiotiques. En général, il semble qu’en dépit de tous les efforts et études effectués sur les MF, il y ait encore une lacune dans les connaissances humaines, et donc d’autres expériences sont nécessaires.
Contributions d’auteur
MS, SK, HT et SL ont conçu le concept. MS, SK, HT et LOS ont fourni un aperçu et rédigé le manuscrit et produit les modèles graphiques et les tableaux. SK, MJ, MI, SL et RDM ont édité le manuscrit. MS, MI et RDM ont édité et formaté les références manuscrites. MS, MI et SL ont révisé et répondu aux commentaires des examinateurs. Tous les auteurs ont lu et accepté la version publiée du manuscrit.
Financement
Cette étude collaborative a été réalisée avec le soutien du Sichuan Landscape and Recreation Research Center, Sichuan, Chine (no de subvention, JGYQ2019025) ; le Département des sciences et technologies de Chengdu, Sichuan, Chine (numéro de subvention, 2019-YFYF-00040-SN) ; et le Plan de soutien double pour la construction disciplinaire de l’Université agricole du Sichuan (subventions nos 1921993553 et 2021993427).
Remerciements
Nous exprimons notre gratitude aux femmes scientifiques – Un programme du Département de technologie scientifique (SR / WOS-A / LS-17/2017) à S. Kataria. Nous remercions également les trois relecteurs anonymes pour leurs commentaires et suggestions constructifs sur cet article.
Les conflits d’intérêts
Les auteurs ne déclarent aucun conflit d’intérêt.
Les références
- Kataria, S.; Jain, M. Magnetopriming atténue les effets néfastes des stress abiotiques chez les plantes. Dans Tolérance des plantes au stress environnemental , 1ère éd.; Rôle des, phytoprotecteurs ; Hasanuzzaman, M., Fujita, M., Oku, H., Islam, TM, Eds. ; CRC Press : Boca Raton, Floride, États-Unis, 2019 ; p. 427–442. [ Google Scholar ] [ CrossRef ]
- Waqas, M.; Korres, NE ; Khan, MD; Nizami, A.-S. ; Deeba, F.; Ali, je. ; Hussain, H. Avancées dans le concept et les méthodes d’amorçage des semences. Dans l’amorçage et le prétraitement des semences et des semis ; Hasanuzzaman, M., Fotopoulos, V., Eds. ; Springer : Berlin/Heidelberg, Allemagne, 2019 ; p. 11–41. [ Google Scholar ] [ CrossRef ]
- Athari Nia, M.; Noori, M.; Ghanati, F. Effet du champ magnétique statique sur certaines caractéristiques physiologiques et biochimiques de Cicer arietinum en phase de croissance végétative. Pajouhesh Sazandegi 2008 , 21 , 62–68. [ Google Scholar ]
- Zhadin, MN Revue de la littérature russe sur l’action biologique des champs magnétiques CC et CA à basse fréquence. Bioélectromagnétique 2001 , 22 , 27–45. [ Google Scholar ] [ CrossRef ]
- Kordas, L. L’effet du champ magnétique sur la croissance, le développement et le rendement du blé de printemps. Pol. J. Environ. Étalon. 2002 , 11 , 527–530. [ Google Scholar ]
- Mahajan, TS ; Pandey, OP Modèle de temps magnétique à la germination hors saison. Int. Agrophys 2014 , 28 , 57–62. [ Google Scholar ] [ CrossRef ]
- Eftimiadou, A.; Katsenios, N.; Karkanis, A.; Papastylianou, P.; Triantafyllidis, V.; Travlos, I. ; Bilalis, DJ Effets du traitement électromagnétique pulsé avant le semis de semences de tomates sur la croissance, le rendement et la teneur en lycopène. Sci. Monde J. 2014 , 1–6. [ Google Scholar ] [ CrossRef ]
- Menegatti, R.D. ; de Oliveira, LO; da Costa, Á.VL; Braga, EJB; Bianchi, VJ Champ magnétique et acide gibbérélique comme traitements de pré-germination des graines de fruits de la passion. Ciência Agrícola Rio Largo 2019 , 17 , 15–22. [ Google Scholar ] [ CrossRef ]
- Abdolmaleki, P.; Ghanati, F.; Sahebjamei, H.; Sarvestani, AS Activité peroxydase, lignification et promotion de la mort cellulaire dans les cellules de tabac exposées à un champ magnétique statique. Environnementaliste 2007 , 27 , 435–440. [ Google Scholar ] [ CrossRef ]
- Ijaz, B.; Jatoi, SA; Ahmad, D.; Masood, MS ; Siddiqui, SU Changements dans le comportement de germination des graines de blé exposées à un champ magnétique et à de l’eau structurée magnétiquement. Afr. J. Biotechnol. 2012 , 11 , 3575–3585. [ Google Scholar ] [ CrossRef ]
- Aladjadjiyan, A. Etude de l’influence du champ magnétique sur certaines caractéristiques biologiques de Zea mais. J.Cent. EUR. Agric. 2002 , 3 , 89–94. [ Google Scholar ]
- Belyavskaya, N. Effets biologiques dus à un champ magnétique faible sur les plantes. Adv. Espace Rés. 2004 , 34 , 1566–1574. [ Google Scholar ] [ CrossRef ]
- Da Silva, JAT ; Dobránszki, J. Champs magnétiques : quel est l’impact sur la croissance et le développement des plantes ? Protoplasma 2016 , 253 , 231–248. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Aladjadjiyan, A. L’utilisation de méthodes physiques pour la stimulation de la croissance des plantes en Bulgarie. J.Cent. EUR. Agric. 2007 , 8 , 369–380. Disponible en ligne : https://hrcak.srce.hr/19607 (consulté le 1er février 2020).
- Chen, HH ; Chang, HC; Chen, YK; Pendu, CL ; Lin, SY; Chen, YS Un procédé amélioré pour une nutrition élevée de la production de riz brun germé : Plasma à basse pression. Chimie alimentaire. 2016 , 191 , 120–127. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Shine, M.; Guruprasad, K.; Anand, A. Amélioration de la germination, de la croissance et de la photosynthèse du soja par prétraitement des graines avec un champ magnétique. Bioélectromagnétique 2011 , 32 , 474–484. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Bhardwaj, J.; Anand, A.; Nagarajan, S. Changements biochimiques et biophysiques associés à la magnéto-amorçage dans la germination des graines de concombre. Physique Végétale. Biochimie. 2012 , 57 , 67–73. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Kataria, S.; Baghel, L.; Guruprasad, K. Accélération de la germination et des caractéristiques de croissance précoce du soja et du maïs après prétraitement des semences avec un champ magnétique statique. Int. J. Trop. Agric. 2015 , 33 , 985–992. [ Google Scholar ]
- Anand, A.; Kumari, A.; Thakur, M.; Koul, A. La signalisation du peroxyde d’hydrogène s’intègre aux phytohormones lors de la germination des graines de tomates magnéto-amorcées. Sci. Rép. 2019 , 9 , 1–11. [ Google Scholar ] [ CrossRef ]
- Thomas, S.; Anand, A.; Chinnusamy, V.; Dahuja, A.; Basu, S. Magnetopriming contourne l’effet du stress de salinité sur la germination des graines de pois chiches. Acta Physiol. Usine 2013 , 35 , 3401–3411. [ Google Scholar ] [ CrossRef ]
- Kataria, S.; Baghel, L.; Guruprasad, K. Le prétraitement des semences avec un champ magnétique statique améliore la germination et les caractéristiques de croissance précoce sous stress salin chez le maïs et le soja. Biocatal. Agric. Biotechnol. 2017 , 10 , 83–90. [ Google Scholar ] [ CrossRef ]
- Reine, FG ; Pascual, LA Influence d’un champ magnétique stationnaire sur les relations hydriques dans les graines de laitue. Première partie : Considérations théoriques. Bioélectromagnétique 2001 , 22 , 589–595. [ Google Scholar ] [ CrossRef ]
- Maheshwari, BL; Grewal, HS Traitement magnétique de l’eau d’irrigation : ses effets sur le rendement des cultures maraîchères et la productivité de l’eau. Agric. Gestion de l’eau 2009 , 96 , 1229–1236. [ Google Scholar ] [ CrossRef ]
- Hilal, M.; Shata, S.; Abdel-Dayem, A.; Hilal, M. Application des technologies magnétiques dans l’agriculture du désert : III. Effet de l’eau magnétisée sur le rendement et l’absorption de certains éléments par les agrumes en relation avec la mobilisation des nutriments dans le sol. Egypte. J. Sol Sci. 2002 , 42 , 43–56. [ Google Scholar ]
- Cakmak, T.; Dumlupinar, R.; Erdal, S. Accélération de la germination et de la croissance précoce des semis de blé et de haricot cultivés sous divers champs magnétiques et conditions osmotiques. Bioélectromagnétique 2010 , 31 , 120–129. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Baghel, L.; Kataria, S.; Guruprasad, KN Le traitement par champ magnétique statique des graines améliore le métabolisme du carbone et de l’azote sous stress de salinité dans le soja. Bioélectromagnétique 2016 , 37 , 455–470. [ Google Scholar ] [ CrossRef ]
- Kataria, S.; Baghel, L.; Guruprasad, K. Atténuation des effets néfastes du stress UV ambiant sur la croissance et certains attributs physiologiques potentiels du soja (Glycine max) par prétraitement des semences avec un champ magnétique statique. J. Plant Growth Regul. 2017 , 36 , 550–565. [ Google Scholar ] [ CrossRef ]
- Kataria, S.; Baghel, L.; Jain, M.; Guruprasad, K. Magnetopriming régule le système de défense antioxydant du soja contre le stress salin. Biocatal. Agric. Biotechnol. 2019 , 18 , 101090. [ Google Scholar ] [ CrossRef ]
- Dhawi, F. Pourquoi les champs magnétiques sont-ils utilisés pour améliorer la croissance et la productivité d’une plante ? Annu. Rés. Rév. Biol. 2014 , 886–896. [ Google Scholar ] [ CrossRef ]
- Maffei, ME Effets du champ magnétique sur la croissance, le développement et l’évolution des plantes. Devant. Usine Sci. 2014 , 5 , 445. [ Google Scholar ] [ CrossRef ][ Version verte ]
- Vian, A.; Davies, E.; Gendraud, M.; Bonnet, P. Réponses des plantes aux champs électromagnétiques à haute fréquence. BioMed Res. Int. 2016 , 1830262. [ Google Scholar ] [ CrossRef ][ Version verte ]
- Sahebjamei, H.; Abdolmaleki, P.; Ghanati, F. Effets du champ magnétique sur les activités enzymatiques antioxydantes des cellules de tabac cultivées en suspension. Bioélectromagnétique 2007 , 28 , 42–47. [ Google Scholar ] [ CrossRef ]
- Minorsky, PV Les variations géomagnétiques affectent-elles le fonctionnement des plantes ? J.Atmos. Sol. Terr. Phys. 2007 , 69 , 1770–1774. [ Google Scholar ] [ CrossRef ]
- Galland, P.; Pazur, A. Magnetoreception chez les plantes. J. Plant Res. 2005 , 118 , 371–389. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Occhipinti, A.; De Santis, A.; Maffei, ME La magnétoréception : une étape incontournable pour l’évolution des plantes ? Tendances Plant Sci. 2014 , 19 , 1–4. [ Google Scholar ] [ CrossRef ]
- Vasishth, A.; Nagarajan, S. L’exposition des graines à un champ magnétique statique améliore la germination et les caractéristiques de croissance précoce du pois chiche ( Cicer arietinum L.). Bioélectromagnétique 2008 , 29 , 571–578. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Vasishth, A.; Joshi, DK Caractéristiques de croissance des graines de maïs exposées au champ magnétique. Bioélectromagnétique 2017 , 38 , 151–157. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Poinapen, D.; Brown, DC; Beeharry, GK L’orientation des semences et l’intensité du champ magnétique ont plus d’influence sur les performances des semences de tomates que l’humidité relative et la durée d’exposition à des champs magnétiques statiques non uniformes. J. Plant Physiol. 2013 , 170 , 1251–1258. [ Google Scholar ] [ CrossRef ]
- Konefał-Janocha, M.; Banaś-Ząbczyk, A.; Bester, M.; Bocak, D.; Budzik, S.; Gorny, S.; Larsen, S.; Majchrowski, K.; Cholewa, M. L’effet des champs électromagnétiques stationnaires et variables sur la germination et la croissance précoce du radis ( Raphanus sativus ). Pol. J. Environ. Étalon. 2019 , 28 , 709–715. [ Google Scholar ] [ CrossRef ]
- Florez, M.; Carbonell, MV ; Martínez, E. Exposition de graines de maïs à des champs magnétiques stationnaires : effets sur la germination et la croissance précoce. Environ. Exp. Bot. 2007 , 59 , 68–75. [ Google Scholar ] [ CrossRef ]
- Kavi, P. L’effet du traitement magnétique des graines de soja sur sa capacité d’absorption d’humidité. Sci. Culte. Calcutta 1977 , 405–406. [ Google Scholar ] [ CrossRef ][ Version verte ]
- Phirke, P.; Kubde, A.; Umbarkar, S. L’influence du champ magnétique sur la croissance des plantes. Semence Sci. Technol. 1996 , 24 , 375–392. [ Google Scholar ]
- Martinez, E.; Florez, M.; Carbonell, M. Effet stimulant du traitement magnétique sur la germination des graines de céréales. Int. J. Environ. Agric. Biotechnol. 2017 , 2 , 375–381. [ Google Scholar ] [ CrossRef ]
- Racuciu, M.; Creanga, D.; Horga, I. Croissance des plantes sous l’influence d’un champ magnétique statique. ROM. J.Phys. 2008 , 53 , 353–359. [ Google Scholar ]
- Kataria, S.; Jain, M.; Tripathi, DK ; Singh, VP Implication de la production d’oxyde nitrique dépendante de la nitrate réductase dans la tolérance au sel induite par magnéto-amorçage dans le soja. Physiol. Usine. 2020 , 168 , 422–436. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Ahamed, M.; Elzaawely, A.; Bayoumi, Y. Effet du champ magnétique sur la germination, la croissance et le rendement des graines de poivron ( Capsicum annuum L.). Asian J. Crop. Sci. 2013 , 5 , 286–294. [ Google Scholar ] [ CrossRef ][ Version verte ]
- Hozayn, M.; EL-Mahdy, A.; Zalama, M. Magnéto-amorçage pour améliorer la germination, les attributs des semis et les performances au champ de l’orge ( Hordeum vulgare L.) sous stress de salinité. Moyen-Orient J. Agric. Rés. 2018 , 7 , 1006–1022. [ Google Scholar ]
- Vasishth, A.; Nagarajan, S. Effet sur la germination et les caractéristiques de croissance précoce des graines de tournesol ( Helianthus annuus ) exposées à un champ magnétique statique. J. Plant Physiol. 2010 , 167 , 149-156. [ Google Scholar ] [ CrossRef ]
- Florez, M.; Alvarez, J.; Martinez, E.; Carbonell, V. Le champ magnétique stationnaire stimule la croissance des racines du riz. ROM. Rép. Phys. 2019 , 71 , 713. [ Google Scholar ]
- Mroczek-Zdyrska, M.; Tryniecki, Ł.; Kornarzyński, K.; Pietruszewski, S.; Gagoś, M. Influence de la stimulation du champ magnétique sur la croissance et les paramètres biochimiques de Phaseolus vulgaris L. J. Microbiol. Biotechnol. Sci alimentaire. 2016 , 9 , 548–551. [ Google Scholar ] [ CrossRef ][ Version verte ]
- Ghanati, F.; Abdolmaleki, P.; Vaezzadeh, M.; Rajabbeigi, E.; Yazdani, M. Application du champ magnétique et du fer pour changer les médicaments d’Ocimum Basilicum. Environnement 2007 , 27 , 429–434. [ Google Scholar ] [ CrossRef ]
- Kornarzyński, K.; Dziwulska-Hunek, A.; Kornarzyńska-Gregorowicz, A.; Sujak, A. Effet de la stimulation électromagnétique des graines d’amarante de différentes humidités initiales sur les paramètres de germination et la teneur en pigments photosynthétiques. Sci. Rép. 2018 , 8 , 1–12. [ Google Scholar ] [ CrossRef ]
- Kato, R.; Kamada, H.; Asashima, M. Effets des champs magnétiques élevés et très faibles sur la croissance des racines poilues de Daucus carota et Atropa belladonna. Cellule végétale Physiol. 1989 , 30 , 605–608. [ Google Scholar ] [ CrossRef ]
- Abobatta, WF Aperçu du rôle de la magnétisation de l’eau traitée dans le développement du secteur agricole. Adv. Agric. Technol. Usine Sci. 2019 , 2 , 180023. [ Google Scholar ]
- Mghaiouini, R.; Elaouad, A.; Taimoury, H.; Sabir, I. ; Chibi, F.; Hozayn, M.; Garmim, T.; Nmila, R.; Rchid, H.; Monkade, M. Influence du dispositif électromagnétique Aqua 4D sur la qualité de l’eau et la germination de la laitue ( Lactuca sativa L.). Int. J. Curr. Ing. Technol. 2020 , 19–24. [ Google Scholar ] [ CrossRef ]
- Tai, CY; Wu, C.-K.; Chang, M.-C. Effets du champ magnétique sur la cristallisation de CaCO 3 à l’aide d’aimants permanents. Chim. Ing. Sci. 2008 , 63 , 5606–5612. [ Google Scholar ] [ CrossRef ]
- Kareem, NSA Évaluation des impacts de la magnétisation de l’eau d’irrigation sur l’amélioration du rendement et de la productivité de l’eau pour certaines cultures. J. Agric. Sci. Technol. A 2018 , 8 , 271–283. [ Google Scholar ] [ CrossRef ][ Version verte ]
- Hirota, N.; Nakagawa, J.; Kitazawa, K. Effets d’un champ magnétique sur la germination des plantes. J. Appl. Phys. 1999 , 85 , 5717–5719. [ Google Scholar ] [ CrossRef ]
- Fernandez, L.; Teran, Z.; Leon, M. L’effet de l’eau d’irrigation traitée magnétiquement sur la qualité des plants d’oignon cultivés en zéoponie. Cultiv. Trop. 1996 , 17 , 55–59. [ Google Scholar ]
- Lin, je.; Yotvat, J. Exposition de l’eau d’irrigation et de l’eau potable à un champ magnétique avec une puissance et une direction contrôlées. J. Magn. Magn. Mater. 1990 , 83 , 525-526. [ Google Scholar ] [ CrossRef ]
- Taimourya, H.; Oussible, M.; Baamal, L.; Harif, A.; Zaid, E.; Guedira, A.; Smouni, A. Le traitement magnétique du milieu de culture améliore la croissance et l’absorption de minéraux de la fraise ( Fragaria × ananassa Duch.) et de la tomate ( Solanum lycopersicum ) dans des conditions de carence en Fe. Int. J. Sci. Ing. Rés. 2017 , 8 , 1414–1436. [ Google Scholar ]
- Noran, R.; Shani, U. ; Lin, I. L’effet de l’irrigation avec de l’eau traitée magnétiquement sur la translocation des minéraux dans le sol. Magn. Électr. Septembre 1996 , 7 , 109–122. [ Google Scholar ] [ CrossRef ]
- Hasan, MM; Alharby, HF ; Hajar, AS ; Hakeem, KR ; Alzahrani, Y. L’effet de l’eau magnétisée sur la croissance et les conditions physiologiques des espèces de Moringa sous stress hydrique. Pol. J. Environ. Étalon. 2019 , 28 , 1145–1155. [ Google Scholar ] [ CrossRef ]
- Migahid, M.; Elghobashy, R.; Bidak, L.; Amin, A. L’amorçage des graines de Silybum marianum (L.) Gaertn avec H 2 O 2 et un champ magnétique améliore le stress de l’eau de mer. Heliyon 2019 , 5 , e01886. [ Google Scholar ] [ CrossRef ] [ PubMed ][ Version verte ]
- Bagherifard, A.; Ghasemnezhad, A. Effet de l’eau salée magnétique sur certaines caractéristiques morphologiques et biochimiques des feuilles d’artichaut ( Cynara scolymus L.). J. Med. Plantes Sous-Prod. 2014 , 3 , 161–170. [ Google Scholar ]
- Reine, FG ; Pascual, LA ; Fundora, IA Influence d’un champ magnétique stationnaire sur les relations hydriques dans les graines de laitue. Partie II : Résultats expérimentaux. Bioélectromagnétique 2001 , 22 , 596–602. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Azimi, N.; Majd, A.; Nejadsattari, T.; Ghanati, F.; Arbabian, S. Effets de l’eau traitée magnétiquement sur les caractéristiques physiologiques de Lens culinaris L. Iran. J. Sci. Technol. Trans. Sci. 2018 , 42 , 331–337. [ Google Scholar ] [ CrossRef ]
- Ul Haq, Z.; Iqbal, M.; Jamil, Y.; Anwar, H.; Younis, A.; Arif, M.; Farid, MZ ; Hussain, F. Effet d’irrigation de l’eau traitée magnétiquement sur la germination des graines de navet, la croissance des semis et les activités enzymatiques. Inf. Processus. Agric. 2016 , 3 , 99–106. [ Google Scholar ] [ CrossRef ][ Version verte ]
- Shahin, M.; Mashhour, A.; Abd-Elhady, E. Effet de l’eau d’irrigation et des graines magnétisées sur certaines propriétés de l’eau, les paramètres de croissance et la productivité des plants de concombre. Courant. Sci. Int. 2016 , 5 , 152–164. [ Google Scholar ]
- Shabrangi, A.; Majd, A.; Sheidai, M. Effets des champs électromagnétiques de fréquence extrêmement basse sur la croissance, la cytogénétique, la teneur en protéines et le système antioxydant de Zea mays L. Afr. J. Biotechnol. 2011 , 10 , 9362–9369. [ Google Scholar ] [ CrossRef ][ Version verte ]
- Hajnorouzi, A.; Vaezzadeh, M.; Ghanati, F.; Nahidian, B. Promotion de la croissance et diminution du stress oxydatif chez les semis de maïs par une combinaison de champs géomagnétiques et électromagnétiques faibles. J. Plant Physiol. 2011 , 168 , 1123–1128. [ Google Scholar ] [ CrossRef ]
- Bradford, MM Une méthode rapide et sensible pour la quantification de microgrammes de protéines utilisant le principe de la liaison protéine-colorant. Anal. Biochimie. 1976 , 72 , 248–254. [ Google Scholar ] [ CrossRef ]
- Clark, G. Procédures de coloration , 4e éd. ; Williams & Wilkins Press : Baltimore/Londres, Royaume-Uni, 1981 ; p. 512. [ Google Scholar ]
- Repacholi, MH; Greenebaum, B. Interaction des champs électriques et magnétiques statiques et à très basse fréquence avec les systèmes vivants : effets sur la santé et besoins de recherche. Bioélectromagnétique 1999 , 20 , 133–160. [ Google Scholar ] [ CrossRef ]
- Greenebaum, B.; Barnes, F. Bioingénierie et aspects biophysiques des champs électromagnétiques , 4e éd.; CRD Press : Boca Raton, Floride, États-Unis, 2018 ; p. 106–230. [ Google Scholar ] [ CrossRef ]
- Mohammadi, F.; Ghanati, F.; Sharifi, M.; Chashmi, NA Sur le mécanisme du contrôle du cycle cellulaire des cellules de tabac cultivées en suspension après exposition à un champ magnétique statique. Usine Sci. 2018 , 277 , 139–144. [ Google Scholar ] [ CrossRef ]
- Vert, LM ; Miller, AB; Agnew, DA; Greenberg, ML; Li, J.; Villeneuve, PJ; Tibshirani, R. Leucémie infantile et surveillance personnelle des expositions résidentielles aux champs électriques et magnétiques en Ontario, Canada. Le cancer provoque le contrôle. 1999 , 10 , 233–243. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Jouni, FJ; Abdolmaleki, P.; Ghanati, F. Stress oxydatif chez la fève ( Vicia faba L.) induit par un champ magnétique statique sous radioactivité naturelle. Mutat. Rés. Genet. Toxicol. Environ. Mutagène. 2012 , 741 , 116–121. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Shine, M.; Guruprasad, K. Impact de l’exposition au champ magnétique avant le semis des graines à un champ magnétique stationnaire sur la croissance, les espèces réactives de l’oxygène et la photosynthèse du maïs dans des conditions de terrain. Acta Physiol. Usine 2012 , 34 , 255–265. [ Google Scholar ] [ CrossRef ]
- El-Yazied, A.; Shalaby, O.; El-Gizawy, A.; Khalf, S.; El-Satar, A. Effet du champ magnétique sur la germination des graines et la croissance des plants de tomates. Confiture. Sci. 2011 , 7 , 306–312. [ Google Scholar ] [ CrossRef ]
- Shabrangi, A.; Majd, A. Effet des champs magnétiques sur la croissance et les systèmes antioxydants dans les plantes agricoles. Dans Actes du symposium sur les progrès de la recherche électromagnétique (PIERS), Pékin, Chine, 23-27 mars 2009 ; p. 23–27. [ Google Scholar ]
- Çelik, Ö.; Büyükuslu, N.; Atak, Ç.; Rzakoulieva, A. Effets du champ magnétique sur l’activité de la superoxyde dismutase et de la catalase dans Glycine max (L.) Merr. Racines. Pol. J. Environ. Étalon. 2009 , 18 , 175–182. [ Google Scholar ]
- Roux, D.; Vian, A.; Girard, S.; Bonnet, P.; Paladian, F.; Davies, E.; Ledoigt, G. Champ électromagnétique haute fréquence (900 MHz) et faible amplitude (5 V m−1) : un véritable stimulus environnemental qui affecte la transcription, la traduction, le calcium et la charge énergétique de la tomate. Planta 2008 , 227 , 883–891. [ Google Scholar ] [ CrossRef ]
- Chen, Y.-P. ; Li, R.; Lui, J.-M. Le champ magnétique peut atténuer l’effet toxicologique induit par le cadmium dans les semis de haricot mungo. Écotoxicologie 2011 , 20 , 760–769. [ Google Scholar ] [ CrossRef ]
- Baghel, L.; Kataria, S.; Jain, M. Atténuation des effets néfastes du stress salin sur la germination, la croissance, l’efficacité photosynthétique et le rendement du maïs ( Zea mays L.) par magnéto-amorçage. Acta Agrobot. 2019 , 72 , 1–16. [ Google Scholar ] [ CrossRef ][ Version verte ]
- Baghel, L.; Kataria, S.; Guruprasad, K. Effet du prétraitement par champ magnétique statique sur la croissance, les performances photosynthétiques et le rendement du soja sous stress hydrique. Photosynthetica 2018 , 56 , 718–730. [ Google Scholar ] [ CrossRef ]
- Anand, A.; Nagarajan, S.; Verma, A.; Joshi, D.; Pathak, P.; Bhardwaj, J. Le prétraitement des semences avec un champ magnétique statique améliore le stress hydrique du sol chez les semis de maïs ( Zea mays L.). Indian J. Biochem. Biol. 2012 , 49 , 63–70. [ Google Scholar ]
- Kataria, S.; Rastogi, A.; Bele, A.; Jain, M. Rôle de l’oxyde nitrique et des espèces réactives de l’oxygène dans la tolérance induite par le prétraitement par champ magnétique statique au stress UV-B ambiant dans le soja. Physiol. Mol. Biol. Usine 2020 , 1–15. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Santos, LO ; Deamici, KM ; Menestrino, C.-B.; Garda-Buffon, J.; Costa, JAV Traitement magnétique des microalgues pour une meilleure formation des produits. Monde J. Microb. Biotechnol. 2017 , 33 , 169. [ Google Scholar ] [ CrossRef ]
- Petit, DP ; Hüner, NP; Wan, W. Effet des champs magnétiques statiques sur la croissance, la photosynthèse et l’ultrastructure des microalgues Chlorella kessleri. Bioélectromagnétique 2012 , 33 , 298–308. [ Google Scholar ] [ CrossRef ]
- Beruto, DT ; Lagazzo, A.; Frumento, D.; Converti, A. Modèle cinétique de croissance de Chlorella vulgaris avec et sans champs électromagnétiques à fréquence extrêmement basse (EM-ELF). J. Biotechnol. 2014 , 169 , 9–14. [ Google Scholar ] [ CrossRef ]
- Wang, HY ; Zeng, XB; Guo, SY ; Li, ZT Effets du champ magnétique sur le système de défense antioxydant de la Chlorella Vulgaris cultivée en recirculation. Bioélectromagnétique 2008 , 29 , 39–46. [ Google Scholar ] [ CrossRef ]
- Tu, R.; Jin, W.; Xi, T.; Yang, Q.; Han, S.-F. ; Abomohra, AE-F. Effet du champ magnétique statique sur la production d’oxygène de Scenedesmus obliquus cultivé dans les eaux usées municipales. Eau Rés. 2015 , 86 , 132–138. [ Google Scholar ] [ CrossRef ]
- Luna, LG ; Menendez, J.; Álvarez, I. ; Flores, I. Effecto de diferentes protocolos de aplicación de un campo magnético (0.03 T) sobre el crecimiento, viabilidad y composición pigmentaria de Haematococcus pluvialis Flotow en suficiencia y ausencia de nitrógeno. Biotechnologie. Végé. 2009 , 9 , 105–117. [ Google Scholar ]
- Chu, F.-J.; Wan, T.-J. ; Pai, T.-Y. ; Lin, H.-W.; Liu, S.-H.; Huang, C.-F. Utilisation de champs magnétiques et de concentration de nitrate pour optimiser la croissance et le rendement lipidique de Nannochloropsis oculata . J. Environ. Géré. 2020 , 253 , 109680. [ Google Scholar ] [ CrossRef ]
- Deamici, KM ; Cardias, BB ; Costa, JAV ; Santos, LO Champs magnétiques statiques dans la culture de Chlorella fusca : effets biologiques sur la croissance et la composition de la biomasse. Processus Biochem. 2016 , 51 , 912–916. [ Google Scholar ] [ CrossRef ]
- Deamici, KM ; Santos, LO ; Costa, JAV Utilisation de champs magnétiques statiques pour augmenter la biofixation du CO2 par la microalgue Chlorella Fusca . Bioressource. Technol. 2019 , 276 , 103–109. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Deamici, KM ; Santos, LO ; Costa, JAV Action du champ magnétique sur les cultures extérieures et intérieures de Spiruline : Évaluation de la croissance, de la consommation moyenne et du profil protéique. Bioressource. Technol. 2018 , 249 , 168–174. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Deamici, KM ; Cuellar-Bermudez, SP ; Muylaert, K.; Santos, LO ; Costa, JAV Modifications du rendement quantique dues à l’action des champs magnétiques statiques sur Arthrospira platensis SAG 21.99 : Évaluation de l’activité du photosystème. Bioressource. Technol. 2019 , 292 , 121945. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Han, S.; Jin, W.; Chen, Y.; Tu, R.; Abomohra, AE-F. Amélioration de la production de lipides de Chlorella pyrenoidosa cultivée dans les eaux usées municipales par traitement magnétique. Appl. Biochimie. Biotechnol. 2016 , 180 , 1043–1055. [ Google Scholar ] [ CrossRef ]
- Huo, S.; Chen, X.; Zhu, F.; Zhang, W.; Chen, D.; Jin, N.; Cobb, K.; Cheng, Y.; Wang, L.; Ruan, R. Intervention de champ magnétique sur la croissance de la microalgue filamenteuse Tribonema sp. dans les eaux usées d’amidon pour la production de biomasse algale et l’élimination des nutriments : influence de la température ambiante et de la stratégie opérationnelle. Bioressource. Technol. 2020 , 303 , 122884. [ Google Scholar ] [ CrossRef ]
- Bauer, LM; Costa, JAV ; da Rosa, APC; Santos, LO Stimulation de la croissance et synthèse de lipides, pigments et antioxydants avec des champs magnétiques dans les cultures de Chlorella kessleri . Bioressource. Technol. 2017 , 244 , 1425–1432. [ Google Scholar ] [ CrossRef ]
- Dodson, Californie ; Horé, PJ ; Wallace, MI Un sens radical de l’orientation : signalisation et mécanisme dans la magnétoréception cryptochrome. Tendances Biochem. Sci. 2013 , 38 , 435–446. [ Google Scholar ] [ CrossRef ]
- Shokrollahi, S.; Ghanati, F.; Sajedi, RH; Sharifi, M. Rôle possible des protéines contenant du fer dans les réponses physiologiques du soja au champ magnétique statique. J. Plant Physiol. 2018 , 226 , 163–171. [ Google Scholar ] [ CrossRef ]
- Qin, L.; Wang, M.; Chen, L.; Liang, X.; Wu, Z.; Lin, Z.; Zuo, J.; Feng, X.; Zhao, J.; Liao, H. Biosynthèse des grappes Fe – S du soja régulée par la fluctuation externe du fer ou du phosphate. Plant Cell Rep. 2015 , 34 , 411–424. [ Google Scholar ] [ CrossRef ]
- Baronia, C.; Chandra, A. Changements dans les attributs biochimiques du siratro causés par l’exposition à l’énergie des micro-ondes. Gestion de gamme Agrofor. 2007 , 28 , 251–252. [ Google Scholar ]
- Mcclean, RG; Schofield, MA; Kean, WF; Sommer, CV; Robertson, DP ; Toth, D.; Gajdardziska-Josifovska, M. Minéraux de fer botaniques : corrélation entre la structure des nanocristaux et les modes d’auto-assemblage biologique. EUR. J. Minéral. 2001 , 13 , 1235–1242. [ Google Scholar ] [ CrossRef ]
- Rajabbeigi, E.; Ghanati, F.; Abdolmaleki, P.; Payez, A. Capacité antioxydante des cellules de persil ( Petroselinum crispum L.) en relation avec les niveaux de ferritine induits par le fer et le champ magnétique statique. Électromagn. Biol. Méd. 2013 , 32 , 430–441. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Ghanati, F.; Payez, A. Fer biofortification et activation du système antioxydant du blé par champ magnétique statique. L’Iran. J. Sci. Technol. A 2015 , 39 , 355–360. [ Google Scholar ] [ CrossRef ]
- Haghighat, N.; Abdolmaleki, P.; Ghanati, F.; Behmanesh, M.; Payez, A. Modification de la catalase et de la MAPK chez Vicia faba cultivée dans un sol à radioactivité naturelle élevée et traitée avec un champ magnétique statique. J. Plant Physiol. 2014 , 171 , 99–103. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Scaiano, J.; Cozens, FL; McLean, J. Modèle pour la rationalisation des effets du champ magnétique in vivo. Application du mécanisme des paires de radicaux aux systèmes biologiques. Photochem. Photobiol. 1994 , 59 , 585–589. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Yaycili, O.; Alikamanoglu, S. L’effet du champ magnétique sur les cultures de tissus de Paulownia. Org. de tissus de cellules végétales. 2005 , 83 , 109–114. [ Google Scholar ] [ CrossRef ]
- Vasishth, A.; Nagarajan, S. Caractérisation de la distribution de l’eau et des activités des enzymes pendant la germination dans des graines de maïs exposées magnétiquement ( Zea mays L.). Indian J. Biochem. Biophys. 2010 , 1 , 311–318. Disponible en ligne : http://nopr.niscair.res.in/handle/123456789/10526 (consulté le 1er février 2020).
- Bébé, SM ; Narayanaswamy, GK ; Anand, A. Production de radicaux superoxydes et indice de performance du photosystème II dans les feuilles de graines de soja magnéto-amorcées. Signal de l’usine. Comportement 2011 , 6 , 1635–1637. [ Google Scholar ] [ CrossRef ][ Version verte ]
- Bahadir, A.; Beyaz, R.; Yildiz, M. Effet du champ magnétique sur la croissance in vitro des semis et la régénération des pousses à partir d’explants de nœuds de cotylédons de Boiss. Bioélectromagnétique 2018 , 39 , 547–555. [ Google Scholar ] [ CrossRef ]
- Zhang, X.; Yarema, K.; Xu, A. Impact du champ magnétique statique (SMF) sur les micro-organismes, les plantes et les animaux. Biol. Eff. Statique Magn. Champs 2017 , 1 , 133–172. [ Google Scholar ] [ CrossRef ]
- De Souza, A.; García, D.; Sueiro, L.; Gilart, F. Amélioration de la germination, de la croissance et du rendement des graines d’oignons par des champs magnétiques non uniformes à très basse fréquence. Sci. Hortique. 2014 , 176 , 63–69. [ Google Scholar ] [ CrossRef ]
© 2020 par les auteurs. Licencié MDPI, Bâle, Suisse. Cet article est un article en libre accès distribué selon les termes et conditions de la licence Creative Commons Attribution (CC BY) ( http://creativecommons.org/licenses/by/4.0/ ).